Archives
- 2026-08
- 2026-07
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-07
-
Despite all efforts for development of
2022-01-03

Despite all efforts for development of reliable laboratory methods, according to WHO/UNAIDS technical update on HIV incidence assays, none of the assays evaluated by Consortium for the Evaluation and Performance of HIV Incidence Assays (CEPHIA) have completely met the criteria for an incidence assay
-
br Materials and methods br Results br Discussion
2022-01-03
Materials and methods Results Discussion In the absence of vascularization phenomena, endothelial cells (ECs) are among the most quiescent cells of the body [27]. ECs proliferation is significant during embryogenesis, as the vasculature is being formed, while in the adult life takes place o
-
The VirScan assay has several advantages
2022-01-03
The VirScan assay has several advantages over alternative multiplex serological assays for peptide discovery. These advantages include quantitative assessment of antibody binding to peptides that span all open reading frames in the HIV genome, including both structural and regulatory proteins; repre
-
The stable association of Hat complexes with histones H
2022-01-03
The stable association of Hat1 complexes with histones H3 and H4 raises a number of interesting issues. The first is the question of how Hat1 remains stably associated with histones H3 and H4. While the simple answer is that Hat1 remains bound to histones because of its association with histone chap
-
Furthermore Rosethorne and Charlton Panula et al Reher
2022-01-03
Furthermore, Rosethorne and Charlton (Panula et al., 2015, Reher et al., 2012, Rosethorne and Charlton, 2011) expressed the human H4R in an osteosarcoma cell line and found that with respect to [35S]GTPγS binding, JNJ-7777120 acted as a neutral antagonist or very weak inverse agonist, a finding whic
-
In order to estimate the influence of mitochondrial
2022-01-03
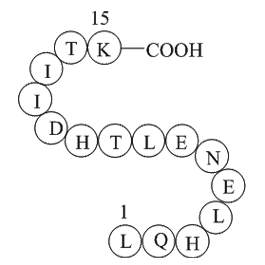
In order to estimate the influence of mitochondrial hypo-polarization on generated OMP, IMP was changed from the normal value of Δψi = −140 mV (Fig. 4A, D, G) to Δψi = −100 mV (Fig. 4B, E, H), resulting in lower magnitudes of calculated OMP for hypo-polarized SM-164 (Fig. 4B). Similarly, a remarkab
-
Compound containing dimethylglutarimide P cap P propyl group
2021-12-31
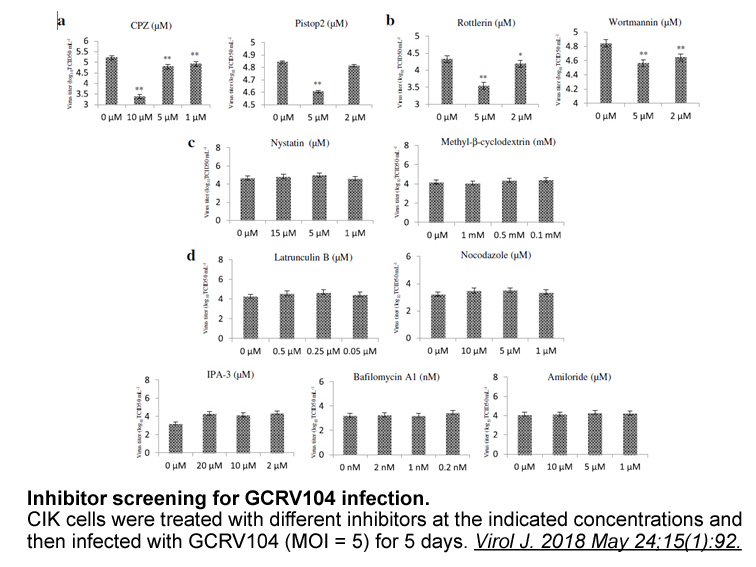
Compound containing, dimethylglutarimide P-cap, P propyl group, α-methylbenzyl urea resulted in inhibitor with much improved binding (=0.064μM) and EC=0.3μM. This was an aza-peptide analog with an EC comparable to our first generation clinical candidate . Analog was evaluated for its selectivity aga
-
Aspartame l aspartyl l phenylalanine methylester is a
2021-12-31
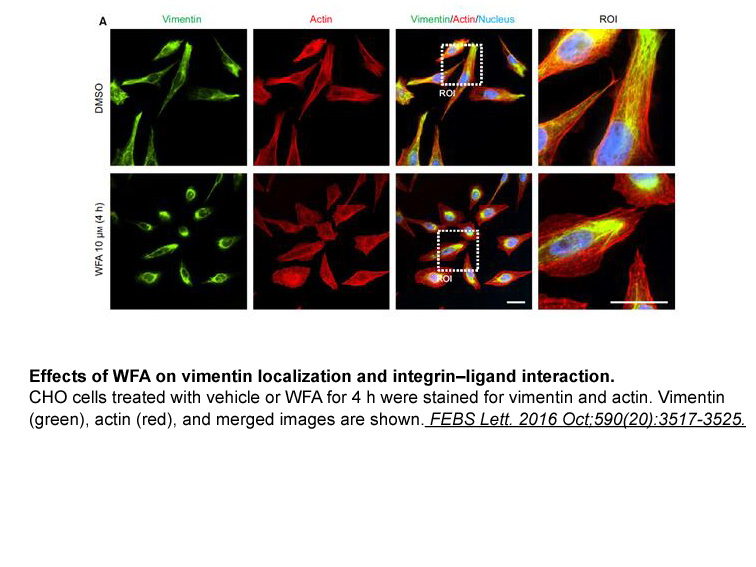
Aspartame (l-aspartyl l-phenylalanine methylester) is a dipeptide artificial sweetener that is widely used as a non-nutritive sweetener in various food products [1]. The increased market of dietary products and the development of new synthetic sweetening compounds have not been sufficiently explored
-
Both plant hormones and some second
2021-12-31
Both plant hormones and some second messengers are involved in Fe-deficiency-related responses in plants (Kobayashi and Nishizawa, 2012). Among these second messengers, nitric oxide (NO) accumulation was induced by Fe deficiencies in tomato and Arabidopsis roots, and it acts as the downstream signal
-
A motivation for this study is the
2021-12-31
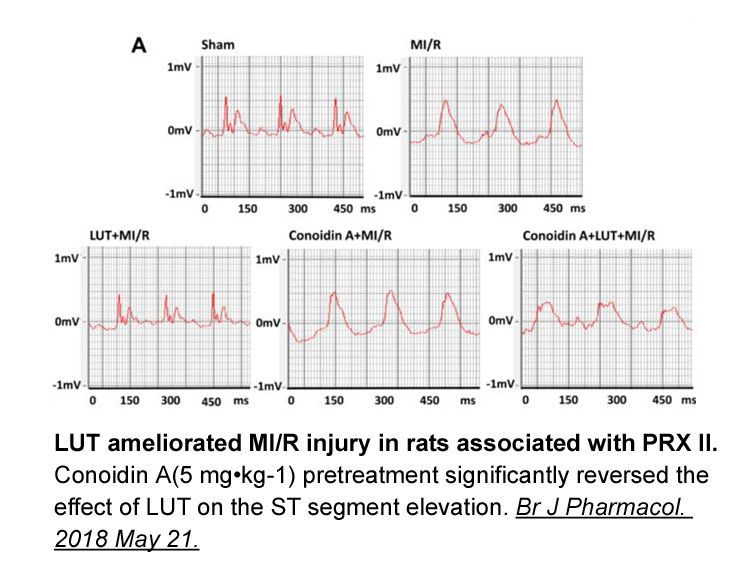
A motivation for this study is the ability of PGD2 to inhibit hair lengthening (). Here we demonstrate that PGD2 also inhibits hair regeneration after wounding. Thus, PGD2 and Gpr44 inhibition of the hair follicle occurs in multiple contexts and may be exploited in future therapies. Pharmaceutical c
-
dna pk Treatment with probiotics can also
2021-12-31
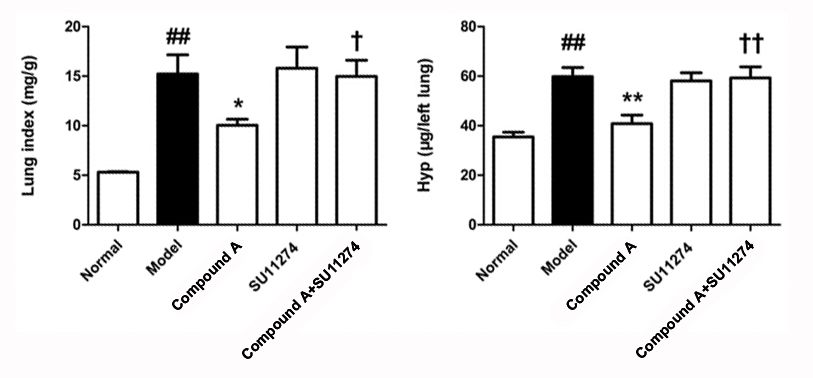
Treatment with probiotics can also be used to alter the microbiota profile of CF patients. One study reports that administration of the probiotic Lactobacillus Reuteri improved digestive health and inflammation [112]. The fecal microbial profile changed, showing a decrease in Proteobacteria and an i
-
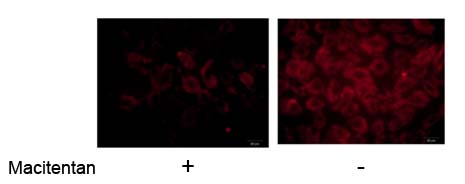
latanoprost In patients with breast cancer ZRANB gene amplif
2021-12-30
In patients with breast cancer, ZRANB1 gene amplification was found in a subset of cases, and ZRANB1 protein levels correlated with EZH2 protein levels and poor survival (both TNBC and non-TNBC). Moreover, unlike normal mammary epithelial cells, all human breast cancer cell lines examined (both TNBC
-
This profound cell type specificity
2021-12-30
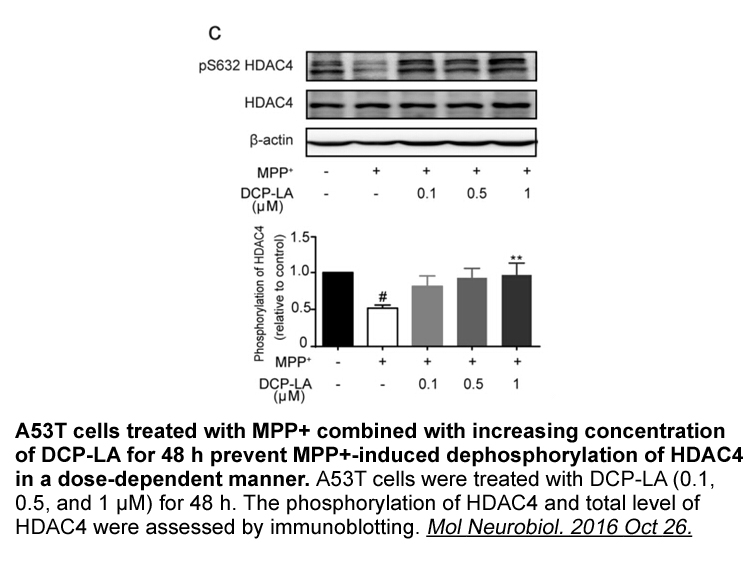
This profound cell-type specificity of G9a/GLP complex modulation of neuronal plasticity is accentuated by studies employing genetic ablation of G9a in dopamine 1 receptor (Drd1)- and dopamine 2 receptor (Drd2)-expressing medium spiny neurons (MSNs) in the nucleus accumbens (NAc). Conditional G9a kn
-
br Acknowledgements This work was supported by grants from t
2021-12-30
Acknowledgements This work was supported by grants from the Agencia Nacional de Promoción Científica y Tecnológica (PICT2013-0495, PICT2016-1821), and from CONICET (PIP 2013-562), as well as from Fundación René Barón and Fundación Williams. Introduction Histamine poisoning, also known as scom
-
p neurotrophin receptor p NTR or CD a
2021-12-30
p75 neurotrophin receptor (p75NTR or CD271), a member of the tumor necrosis factor receptor superfamily, has been implicated in several steps of glioma tumorgenesis [5]. p75NTR mediates glioma invasion and progression through γ-secretase-dependent and -independent mechanisms [6], [7], and its expres
16256 records 555/1084 page Previous Next First page 上5页 551552553554555 下5页 Last page